

Saline Oscillation as a Forge for Coevolution
Why C. Albicans and Humans are The Most Advanced

[Hello again, Reader. Today, I’m taking another detour to share a new addition to my paper on the Saline Oscillation Hypothesis. Recent literature (Song et al., 2025; Barnett et al., 2025), serves as yet another foundational brick in the Redacted Science Framework theory of coevolution of Candida albicans and Hominids. This addition discusses those papers and develops an analogy of the oscillations as a forge for the advancement of the obligate partnership. It has always been a part of us. Every mammal would appear to have a fungus as a symbiote, but that’s not in scope for today. If you have stumbled upon this article and are not familiar with the Redacted Science corpus, this is not actually a bad place to jump in.
This part isn’t new to the paper, but it is important context for the addition:]
Geological and Paleoclimatic Context
2.1 The East African Rift Valley as Cradle of Hominid Evolution
The East African Rift System (EARS) is an active continental rift zone extending over thousands of kilometers from the Red Sea to Mozambique (Chorowicz, 2005). The region between Lake Turkana and Lake Natron is designated the “Cradle of Mankind” based on the density and significance of hominid fossil discoveries, including Australopithecus afarensis (“Lucy,” ~3.2 Ma; Johanson and White, 1979), Homo erectus (KNM-WT 15000 “Turkana Boy,” ~1.6 Ma; Brown et al., 1985; Walker and Leakey, 1993), the Lomekwi stone tools (~3.3 Ma; Harmand et al., 2015), and Paranthropus boisei (“Nutcracker Man,” ~1.75 Ma; Leakey, 1959).
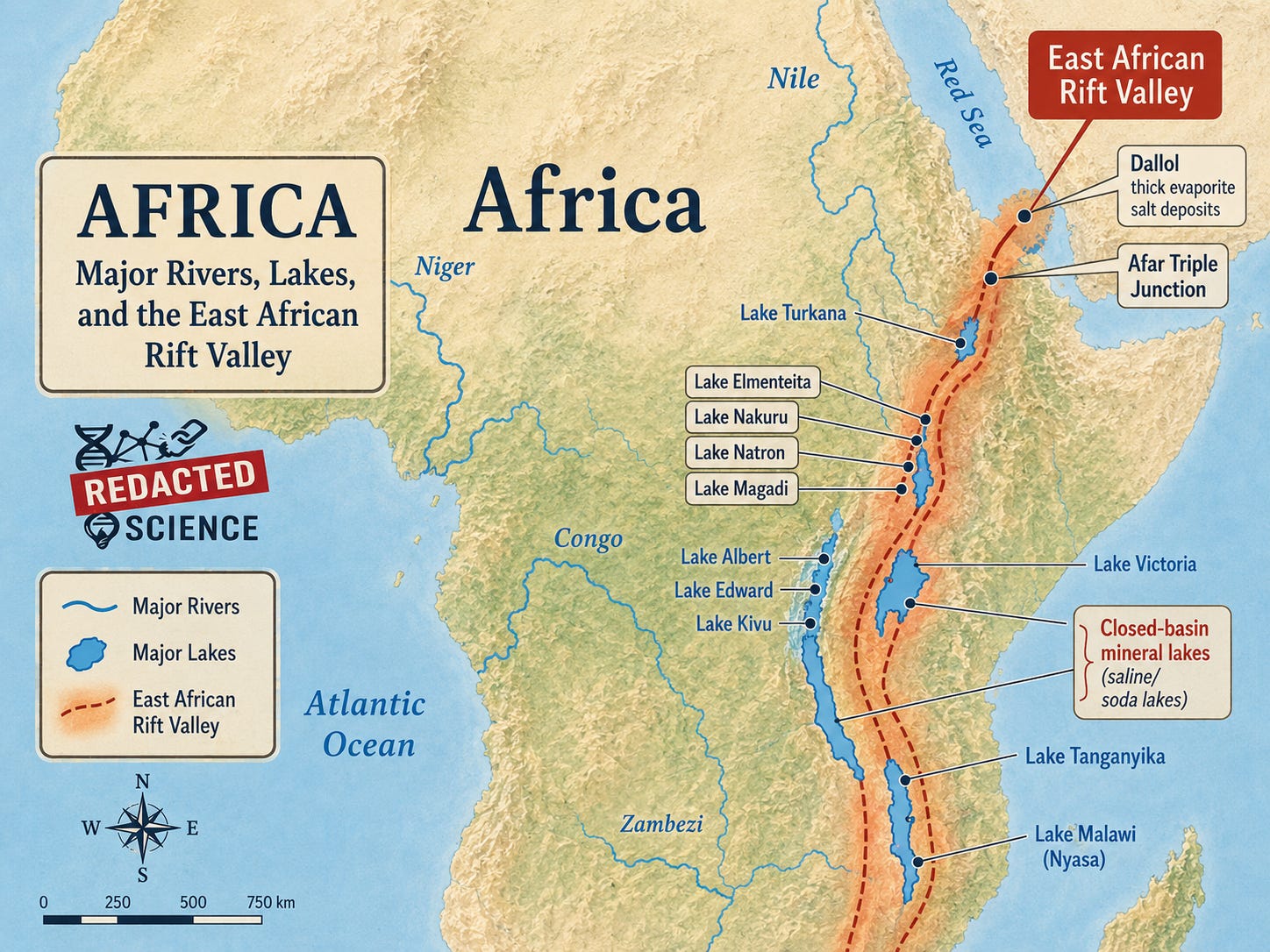
At Dalol, approximately 100 km north of the Afar Triple Junction, the rift floor contains a 5,000-meter-thick layer of evaporite salt deposits accumulated over the past four million years (Ebinger et al., 2000). The Eastern Rift lakes including Turkana, Magadi, Natron, Nakuru, and Elmenteita, are hydrologically closed basins with no outlet to the sea, resulting in high mineral content as evaporation concentrates dissolved salts (Britannica, “East African lakes”). Lake Magadi and Lake Natron are hypersaline soda lakes enriched in Na⁺, K⁺, Cl⁻, CO₃²⁻, and HCO₃⁻ (Deocampo and Renaut, 2022).
2.2 Orbital Forcing, Amplifier Lakes, and Oscillation Architecture
The salinity oscillations central to this hypothesis were not random climate fluctuations. They were driven by predictable variations in Earth’s orbital geometry, amplified by the unusual physical characteristics of rift basin lakes, and structured across at least three nested timescales.
The primary driver is axial precession. Earth’s rotational axis wobbles on a cycle of approximately 19,000 to 23,000 years, commonly averaged to ~21,000 years (Berger and Loutre, 1991). Precession controls the seasonal distribution of solar radiation at tropical latitudes. At precession minima, Northern Hemisphere summer insolation increases, strengthening the African monsoon system, driving rainfall over East Africa, and filling rift basin lakes. At precession maxima, the monsoon weakens, rainfall decreases, and lakes contract (Kutzbach and Street-Perrott, 1985; Trauth et al., 2005). Each precession cycle thus produces one complete wet-dry oscillation in the EARS. Over the 2.7 million years since the onset of intense climate variability, approximately 129 such cycles have occurred. Not all produced equivalent lake-level responses; the amplitude of each cycle depends on a second orbital parameter.
Eccentricity, the degree of circularity of Earth’s orbit, varies on 100,000-year and 400,000-year cycles and modulates the amplitude of precession’s effects (Berger and Loutre, 1991). When eccentricity is high and the orbit more elliptical, precession-driven insolation differences are amplified and the wet-dry swings are extreme. When eccentricity is low, precession effects are damped and climate varies less. This modulation produces what Maslin, Trauth, and colleagues term “variability packets”: concentrated intervals of extreme environmental oscillation separated by periods of relative calm.
Before 2.7 Ma, major wet phases in the EARS appeared approximately every 400,000 years. After 2.7 Ma, East Africa did not oscillate continuously. Instead, the intense variability came in bursts: three windows, each roughly 200,000 years long, separated by calmer intervals of approximately 800,000 years. These windows occurred at 2.7–2.5 Ma, 1.9–1.7 Ma, and 1.1–0.9 Ma (Trauth et al., 2005, 2007; Maslin et al., 2014). Each coincides with a major global climate transition: the onset of Northern Hemisphere glaciation (2.7–2.5 Ma), intensification of the Walker Circulation (1.9–1.7 Ma), and the Mid-Pleistocene Revolution (1.0–0.7 Ma). During these windows, the landscape alternated between large freshwater lakes and extreme drought as rapidly as every 10,000 years, fast enough that a single population lineage would experience the full wet-dry-wet cycle dozens of times within each window (Trauth et al., 2003; Kingston et al., 2007). [Stay with me - coevolution is worth it, I think.]
Within individual precession-driven wet phases, a third timescale operates. Wilson et al. (2014) analyzed oxygen isotope composition and diatom assemblage data from a well-dated diatomite sequence in the Baringo-Bogoria basin and identified millennial-scale cyclicity of 1,400 to 1,700 years between 2.70 and 2.55 Ma, similar in period to late Quaternary Dansgaard-Oeschger events. These sub-oscillations, nested within the larger precession cycles, mean that even during nominally humid phases, lake depth, salinity, and drinking water chemistry fluctuated on timescales of tens of human generations.
The rift basins themselves amplify these climate signals. Trauth et al. (2010) introduced the concept of “amplifier lakes”: tectonic graben morphologies combining high precipitation in elevated catchment areas with extreme evaporation on the valley floor. These basins do not respond proportionally to climate forcing. They respond disproportionately. A moderate shift in the precipitation-evaporation regime produces a dramatic shift in lake level and chemistry, because the rift geometry concentrates the hydrological response (Trauth et al., 2005, 2010). The Eastern Rift lakes, including Turkana, Baringo, Magadi, Natron, and Elmenteita, are hydrologically closed basins with no outlet to the sea, meaning dissolved salts accumulate as evaporation concentrates them (Britannica, “East African lakes”). These are the same basins in which the key hominid fossils were found. [We’re just getting to the good stuff]
Illustration A: Anatomy of a Single Precession Cycle

2.3 Transition Dynamics, Oscillation Count, and the Asymmetric Trigger
The biological significance of the oscillation architecture depends on a critical asymmetry in transition dynamics that has not been previously connected to hominid evolution.
The drying transition is gradual. The sinusoidal character of precession forcing produces intervals of approximately 8,000 years at both the wet and dry extremes during which relatively little change in daily insolation occurs (Maslin & Trauth, 2009), representing stability plateaus during which physiological acclimation is reinforced. Between these plateaus, the wet-to-dry transition unfolds across approximately 14,000 years per cycle as monsoon strength declines progressively, lake level falls, dissolved salts concentrate through evaporation, and vegetation retreats from lakeshore environments. The final desiccation event itself — the contraction from shrinking saline lake to dry basin — takes up to 2,000 years (Bergner et al., 2003; Garcin et al., 2009), nested within the broader drying transition rather than constituting its full duration. Across the full 14,000-year drying period, populations drinking from these water sources experience a slow increase in electrolyte intake spanning dozens of generations, sufficient time for physiological acclimation. Renal sodium handling, blood volume set points, and osmotic equilibria adjust gradually to the rising baseline. [This puts a slow uniform pressure on the population trending A —> B. Your body adapts. The relationship with Candida deepens.]
The freshening transition is not gradual. When monsoon-driven rainfall returns and rift basins fill, lake freshening occurs on timescales far shorter than the drying process. [Boom! Suddenly another pressure happens relatively quickly. Only the strong survive.] A population whose physiology has been calibrated to elevated electrolyte concentrations over generations suddenly encounters freshened water. This is the physiological equivalent of a modern human adapted to normal dietary sodium suddenly drinking distilled water. The result is dilutional hyponatremia and a physiological response analogous to the syndrome of inappropriate antidiuretic hormone secretion (SIADH): water retention, reduced urine output, blood volume expansion, and electrolyte imbalance. The disruption is proportional to the delta between the population’s calibrated salinity and actual intake, not to the absolute salinity level. [I know what it feels like, but that’s another paper]
This asymmetry is the fulcrum of the hypothesis. The slow drying transition produces no acute physiological crisis; populations acclimate. The rapid freshening transition produces a punctuated shock at a specific, predictable point in every oscillation cycle. It is at this point, and only at this point, that the symbiont’s perfusion-management and electrolyte-handling capabilities become decisive. The selective advantage is not continuous. It is periodic, recurring at the freshening edge of every oscillation, and it is acute. The biochemical computer (Craddock, Biochemical Computer), when exposed to rapidly changing external pressures, responds by adjusting its regulatory and chromatin state. Existing epigenetic configurations that worked before get reinforced and passed forward. Recurrent environmental inputs tend to produce consistent transcriptional outputs, while novel conditions may induce transitions into alternative regulatory states, enabling the emergence of new outputs.
The direct sedimentary evidence for the scale of these oscillations comes from the Lake Malawi Drilling Project. Lyons et al. (2015) recovered the first continuous 1.3-million-year record of continental hydroclimate from an African lake interior, documenting 24 distinct lake-level drops exceeding 200 meters, of which 15 were severe events with water levels reduced more than 400 meters below modern. The distribution of these events was not uniform. Before the Mid-Pleistocene Transition (~800 ka), lake levels were generally lower and changed frequently, consistent with a drier baseline climate with rapid oscillations. After the MPT, the lake was commonly deeper and often overflowing, but minimum standing lake level intervals became more prolonged and extreme (Lyons et al., 2015). Johnson et al. (2016) characterized the post-MPT record as dominated by strong 100,000-year eccentricity cycles of temperature and rainfall superimposed on a trend toward progressively wetter conditions.
The Lake Malawi record, while the most continuous available, comes from the southern EARS (10–14° S), approximately 2,000 km south of the Turkana-Baringo-Natron corridor where the key hominid fossils were found. The northern rift basins show stronger precessional control on lake levels (Kingston et al., 2007; Deino et al., 2006) and dried out faster than the rest of Africa (Turkana Basin Institute, 2021), indicating that hominid populations in the cradle region likely experienced more frequent and more intense oscillations than the Malawi record documents.
A conservative accounting of the total oscillation exposure experienced by EARS hominid populations since 2.7 Ma includes: approximately 129 precession-driven cycles (orbital mechanics), concentrated into three variability windows totaling roughly 600,000 years of intense oscillatory pressure, with millennial-scale sub-oscillations numbering in the hundreds nested within those precession cycles. Each cycle, at the freshening transition, presented the selective filter through which symbiont-integrated individuals passed and unintegrated individuals did not.
[Ok, Here is the new part. The Framework only gets stronger from here. When Co-Scientist or Robin/Finch or another such AI service is utilized against the Framework, it will start to become part of the true AI corpus. From there, things will have to change. Do not underestimate AI]
Two independent lines of recent evolutionary biology research reinforce the central claim of this paper: the partnership architecture is a maintained state, not a default. Song et al. (2025) showed that beneficial mutations are far more abundant than Neutral Theory assumed, yet rarely reach fixation because environments shift before selection can complete its work — populations continuously chase moving targets. Barnett et al. (2025) experimentally demonstrated that under cycling selection, lineages evolve heritable localized hypermutability biased toward generating adaptive variation at specific loci. Together, these findings establish that the correct genomic and regulatory architecture can sustain partnership-relevant functions across prolonged environmental oscillation.
This paper proposes a further architectural consequence: the EARS-scenario saline oscillation cycled for hundreds of thousands of years within a range the partnership architecture was selected to track, alternating between freshwater and hypersaline extremes through a structure of sustained selective pressure followed by rapid transition. This cycling regime functions structurally as a forge (Figure A).
A forge transforms metal not in a single step, but through repeated cycles: the metal is heated until it becomes workable, hammered into a new form, then quenched or cooled to lock that form in place before the process begins again. Each cycle does specific work. Heat opens the structure, shaping forces it into a new configuration, and cooling preserves the change so the metal does not simply return to what it was. In this paper, the saline oscillation is analogous to that process: sustained environmental phases do the heating and working, rapid transitions do the quenching, and over many cycles the host-organism partnership is not merely selected, but forged into a deeper, more durable architecture.
The forge metaphor is not novel as evolutionary mechanism — mainstream evolutionary biology already accepts that cycling environments shape genome architecture in ways that stable environments do not, that alternating selective regimes produce more robust adaptation than constant pressure, and that the work done by selection during specific cycle phases shapes the resulting architecture in specific ways.
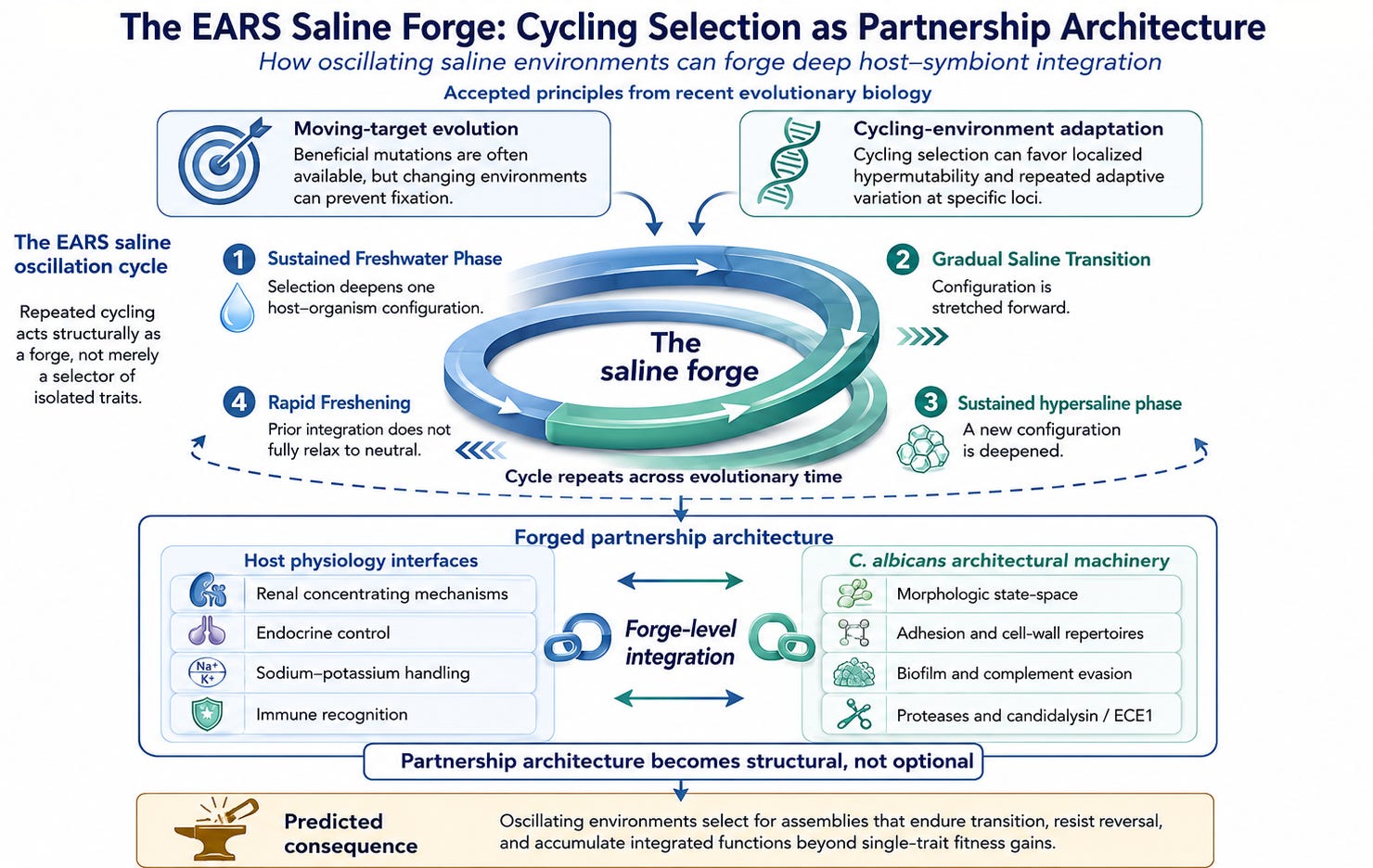
Figure A. Recent evolutionary work supports the principle that cycling environments can shape adaptive architecture differently from stable environments. In the Saline Oscillation Hypothesis, repeated freshwater-to-hypersaline cycling in the East African Rift System functions as a forge: sustained phases deepen host-organism integration, while a gradual salination followed by rapid freshening ratchet configurations forward before prior adaptations can relax to neutrality. Across many cycles, the result is not a detachable opportunistic association but a structurally integrated host-symbiont partnership architecture. The unusually elaborate molecular repertoire of C. albicans is presented as a predicted signature of this forge-level integration rather than merely a retained set of virulence factors.
The framework’s contribution is extending this accepted principle to its full implication for coevolutionary partnerships rather than for single-lineage adaptation. Each EARS oscillation cycle does specific work on the partnership architecture: sustained selection within a configuration deepens the integration; rapid transition locks new configurations in place; the next sustained phase deepens integration of the new configuration without permitting the prior integration to relax to neutral. Over many cycles across hundreds of thousands of years, the partnership architecture becomes forged at the structural level — not merely selected, but shaped by the specific molecular interfaces between host and organism across each cycle’s selective regime. [More specifically, apply pressure in one uniform direction. Reset just within the edge of survivability. Start Over]
Two implications follow directly. First, the partnership architecture present in modern Homo sapiens cannot have been built by substituting C. albicans for an earlier partnership organism at any point in the lineage’s history; forge-level integration requires the same molecular interfaces operating across the cycling history. The pan-mammalian observation that each mammalian lineage carries its own lineage-specific Saccharomycetaceae symbiont (Craddock, Pan-Mammalian) reflects this constraint — each host-organism pairing was forged together from early in the lineage’s evolutionary history. Second, the disruption signatures observed when modern pharmacology removes partnership components (Lionakis et al., 2008; Lionakis section in Kemper et al., 2023) are predicted by forge-level integration: the architecture cannot be unforged without disruption proportional to the depth at which the partnership has been integrated into host physiology. [This partnership is not optional]
The framework’s prediction that the forge process produces deeper architectural integration than ordinary selection has a worked example within the Candida genus itself. C. albicans is documented across the mycology literature as having the most structurally and behaviorally complex molecular repertoire of any pathogenic Candida species: the most elaborate morphological state-space, the largest secreted aspartyl protease family, the most diverse adhesin and immunomodulatory cell wall component repertoires, the most developed biofilm-forming and complement evasion machinery, and the most extensive candidalysin/ECE1 system. Sister species including C. dubliniensis, C. glabrata, C. tropicalis, and others show substantially reduced complements of this architectural machinery. The mainstream framing treats this complexity differential as C. albicans has more virulence factors than its sister species,’ but the framing has a problem the field has not adequately addressed: virulence factors should be costly to maintain when not actively producing pathogenesis, and natural selection should have pruned the elaborate machinery toward whatever subset is consistently advantageous. The persistence and continuous refinement of C. albicans‘s elaborate architecture across evolutionary time is inconsistent with the standard opportunism narrative. The forge model resolves this: C. albicans‘s elaborate machinery is not a collection of retained virulence factors but the partnership architecture that the EARS-scenario forge specifically produced. The complexity gradient across the Candida genus tracks the gradient of coevolutionary exposure intensity — C. albicans was the species in the forge, sister species were not, and the documented architectural complexity differential is what the forge model predicts the outcome should look like.
[Coevolution. Pretty obvious when you look at it like that.]
[I’m not sure what is next. I have several irons in the fire, but I also like just goofing off, so… we’ll see. BTW, I welcome questions] - The Architect
References
Barnett, M., Meister, L., & Rainey, P. B. (2025). Experimental evolution of evolvability. Science. https://doi.org/10.1126/science.adr2756
Berger, A., & Loutre, M. F. (1991). Insolation values for the climate of the last 10 million years. Quaternary Science Reviews, 10(4), 297–317. https://doi.org/10.1016/0277-3791(91)90033-Q
Bergner, A. G. N., Trauth, M. H., & Bookhagen, B. (2003). Magnitude of precipitation/evaporation changes in the Naivasha Basin (Kenya) during the last 150 kyrs. Global and Planetary Change, 36(1–2), 117–135. https://doi.org/10.1016/S0921-8181(02)00178-9
Brown, F., Harris, J., Leakey, R., & Walker, A. (1985). Early Homo erectus skeleton from west Lake Turkana, Kenya. Nature, 316(6031), 788–792. https://doi.org/10.1038/316788a0
Chorowicz, J. (2005). The East African rift system. Journal of African Earth Sciences, 43(1–3), 379–410. https://doi.org/10.1016/j.jafrearsci.2005.07.019
Craddock, J. The Pan-Mammalian Coevolution Hypothesis. Zenodo. https://doi.org/10.5281/zenodo.19643601 [Cited as: Craddock, Pan-Mammalian.]
Deocampo, D. M., & Renaut, R. W. (2022). Mineral precipitation and hydrochemical evolution through evaporitic processes in soda brines (East African Rift Valley). Chemical Geology, 613, Article 121153. https://doi.org/10.1016/j.chemgeo.2022.121153
Ebinger, C. J., et al. (2000). Rift deflection, migration, and propagation: Linkage of the Ethiopian and Eastern rifts, Africa. GSA Bulletin, 112(2), 163–176.
Garcin, Y., Junginger, A., Melnick, D., Olago, D., Strecker, M. R., & Trauth, M. H. (2009). Late Pleistocene–Holocene rise and collapse of Lake Suguta, northern Kenya Rift. Quaternary Science Reviews, 28(9–10), 911–925. https://doi.org/10.1016/j.quascirev.2008.11.001
Harmand, S., et al. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521(7552), 310–315. https://doi.org/10.1038/nature14464
Johanson, D. C., & White, T. D. (1979). A systematic assessment of early African hominids. Science, 203(4378), 321–330.
Kutzbach, J. E., & Street-Perrott, F. A. (1985). Milankovitch forcing of fluctuations in the level of tropical lakes from 18 to 0 kyr BP. Nature, 317(6033), 130–134. https://doi.org/10.1038/317130a0
Leakey, L. S. B. (1959). A new fossil skull from Olduvai. Nature, 184(4685), 491–493.
Maslin, M. A., & Trauth, M. H. (2009). Plio-Pleistocene East African pulsed climate variability and its influence on early human evolution. In F. E. Grine, J. G. Fleagle, & R. E. Leakey (Eds.), The first humans — Origin and early evolution of the genus Homo. Springer. https://doi.org/10.1007/978-1-4020-9980-9_13
Song, S., Chen, P., Shen, X., & Zhang, J. (2025). Adaptive tracking with antagonistic pleiotropy results in seemingly neutral molecular evolution. Nature Ecology & Evolution. https://doi.org/10.1038/s41559-025-02887-1
Trauth, M. H., et al. (2010). Human evolution in a variable environment: The amplifier lakes of Eastern Africa. Quaternary Science Reviews, 29(23–24), 2981–2988. https://doi.org/10.1016/j.quascirev.2010.07.007
Trauth, M. H., Maslin, M. A., Deino, A., & Strecker, M. R. (2005). Late Cenozoic moisture history of East Africa. Science, 309(5743), 2051–2053. https://doi.org/10.1126/science.1112964
Walker, A., & Leakey, R. (1993). The Nariokotome Homo erectus skeleton. Harvard University Press.